走化性と細胞自律性が「意味生成」の入口になる理由
生命の最小単位である細胞は、外部の化学物質を感知して移動し(走化性)、自らの代謝ネットワークを閉ループで維持しながら環境に適応する(細胞自律性)。これらの現象は一見すると純粋な物理化学的プロセスに見えるが、近年の生命科学・認知科学・哲学の交差点において、**「意味を生成する最小のシステム」**として再評価されている。
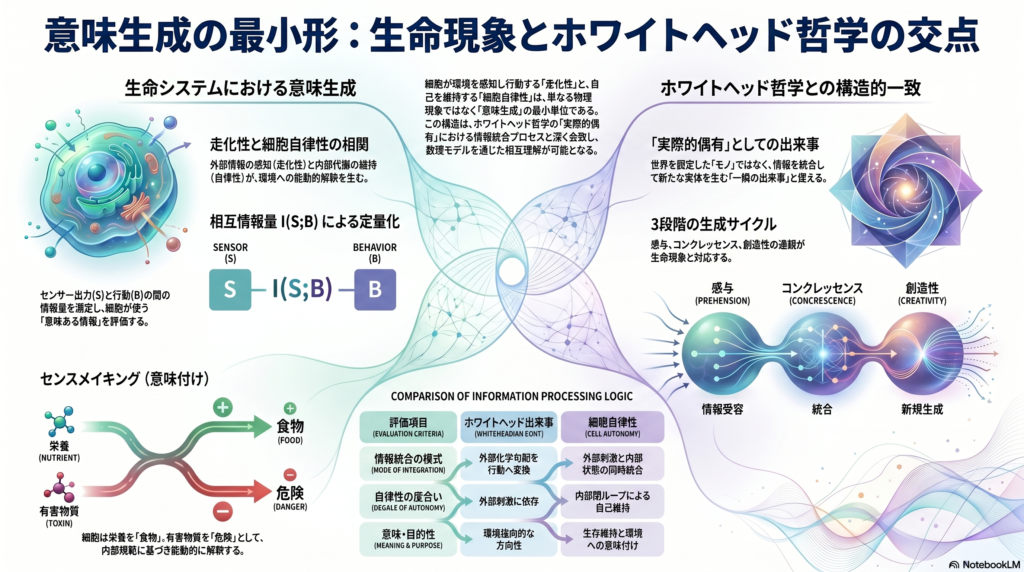
細胞が勾配情報を取り込んで行動に変換するとき、そこには情報の処理と解釈が伴う。この営みは、哲学者アルフレッド・ノース・ホワイトヘッドが20世紀前半に提唱した「実際的偶有(actual occasion)」という概念と驚くほど親和的である。本記事では、走化性と細胞自律性をホワイトヘッド的な「出来事分解」の観点と接続しながら、比較可能な数理モデルへと落とし込む枠組みを丁寧に解説する。
ホワイトヘッドの「実際的偶有」とは何か
プロセス哲学の基本単位としての出来事
ホワイトヘッドの哲学体系、通称「プロセス哲学」では、宇宙を構成する究極の基本単位は**実際的偶有(actual occasion)と呼ばれる。これは、空間に固定した「モノ」ではなく、時間的に離散した一瞬の経験的出来事である。各実際的偶有は、過去に存在したあらゆる情報を「感与(prehension)」**として取り込み、新たな実体へと統合(コンクレッセンス)する過程を担う。
重要なのは、この枠組みが意識を前提としない点だ。ホワイトヘッドにとって、意識的経験は実際的偶有の高次モードに過ぎず、非意識的な物理・生物過程もまた同じ論理構造を持つと考えられた。つまり、細胞レベルの情報統合にも「出来事としての構造」を適用する余地がある。
感与・コンクレッセンス・創造性
実際的偶有の構造を整理すると、次の三つのキーワードが浮かび上がる。
- 感与(Prehension): 過去の情報や周囲の影響を「受け取る」プロセス
- コンクレッセンス(Concrescence): 受け取った情報を統合して新たな実体を生み出すプロセス
- 創造性(Creativity): 各偶有が先行するものをただ反復せず、必ず何らかの新規性を帯びるという原理
この三段階のサイクルは、細胞が化学勾配を感知し(感与)、内部状態に照らして行動を選択し(コンクレッセンス)、次の状態へと移行する(創造性・新規生成)プロセスと構造的に対応している可能性がある。
走化性:意味生成の最小形としての数理モデル
Keller–Segel方程式の骨格
走化性の代表的な数理モデルとして知られるKeller–Segel型の反応拡散方程式は、細胞密度 u(x,t) と誘引物質濃度 c(x,t) の時空間ダイナミクスを記述する。∂t∂u=Du∇2u−χ∇⋅(u∇c)+f(u) ∂t∂c=Dc∇2c+g(u,c)−h(c)
ここで χ(走化感受度)が大きいほど、細胞は濃度勾配 ∇c に強く引き寄せられる。この方程式系は、大腸菌の群れ集合行動やパターン形成実験(Budrene & Berg, 1995)と対応することが知られており、走化性という「環境情報の意味ある利用」を数学的に定式化した先駆的モデルである。
情報理論的視点:細胞が「使える情報」の限界
近年の研究は、走化性を情報理論の枠組みで捉え直す試みへと発展している。Mattinglyら(2021)は、大腸菌が走化性において取得・利用できる情報量にはシャノン情報理論的な限界があり、実際の大腸菌はその理論限界に近い効率で情報を使っていることを示した。
センサー出力 S と行動 B(直進・転向など)の間の相互情報量 I(S;B)I(S;B) I(S;B) を評価することで、細胞がどれだけ「意味のある情報」を行動に反映しているかを定量化できる。I(S;B)=s,b∑p(s,b)logp(s)p(b)p(s,b)
この指標は、ホワイトヘッド的な「感与の深さ」と機能的に対応する可能性がある。どれだけ過去の環境情報を統合して現在の行動を形作るか、という問いを数値として扱えるからだ。
ラン・アンド・タンブル運動と確率過程モデル
大腸菌の運動様式として知られる「ラン(直進)とタンブル(転向)」は、確率過程モデルで自然に表現できる。セルの状態遷移確率が局所的な化学濃度勾配に依存するよう設定すると、勾配上昇方向へのバイアスが生まれ、走化性が創発する。このモデルでは、各タイムステップが「直前の感知情報を統合して次の行動を決定する出来事」として機能し、ホワイトヘッドの実際的偶有の連鎖と類比可能な構造をなす。
細胞自律性:オートポイエーシスと「操作的閉鎖」
マトゥラーナとヴァレラの自己生成理論
HumbertoMaturanaとFrancisco Varelaが提唱したオートポイエーシス(自己生成)理論は、細胞自律性の理論的基盤として広く参照される。この理論によれば、生きている細胞は自身を構成する要素を絶えず自ら再生産し、自己の境界を維持する代謝ネットワークによって成り立っている。代謝が止まれば細胞は死ぬという「危うい非平衡状態」こそが、自律的主体の証しである。
この自己完結的な閉ループ構造は、ホワイトヘッドが各実際的偶有に見た「自己完結的生成(self-creative synthesis)」と親和性が高い。それぞれの出来事が過去から引き継ぎながらも自律的に新たな実体を生み出すように、細胞も環境から材料を取り入れながら自己を絶えず再構成する。
適応性とセンスメイキング
Di Paolo(2009)が導入した**適応性(adaptivity)**の概念は、細胞自律性をさらに豊かにする。細胞が栄養欠乏を「感知」して代謝経路を切り替える行為は、単なる物理化学的反応ではなく、環境の状態に「意味」を付与する行為として解釈できる。栄養源は「食物(ポジティブな価値を持つ対象)」として、有害物質は「危険」として経験される。
このセンスメイキング(意味生成)の視点は、Godfrey-Smith(2016)が最小認知の事例として大腸菌の走化性を挙げたこととも重なる。細胞は刺激に受動的に反応するのではなく、内部の規範(生存・維持)に照らして環境を能動的に解釈し行動する主体であると見なされる。
三者の比較枠組み:走化性・細胞自律性・実際的偶有
| 評価項目 | 走化性 | 細胞自律性 | ホワイトヘッド的出来事 |
|---|---|---|---|
| 情報統合の様式 | 外部化学勾配をセンサーで取得し行動へ変換 | 外部刺激と内部代謝状態を同時統合 | 過去の全情報を「感与」として統合 |
| 自律性の度合い | 外部刺激依存。完全閉ループではない | 内部閉ループで自己維持。環境依存低 | 各偶有は自己完結的な生成単位 |
| 意味・目的性 | 誘引物質へ向かう方向性(環境指向的) | 生存維持が目的。環境を意味づけ | 内的目的(志向性)を各偶有が内包 |
| 時間スケール | 秒〜分単位の応答 | 分〜日単位の恒常性維持 | 極めて瞬間的。次の偶有へ即移行 |
| 代表的モデル | Keller–Segel方程式、確率過程 | 代謝ネットワーク、オートポイエーシス系 | 概念的・定性的(プロセス論的グラフ論) |
この比較表が示すのは、三つの概念が**「情報を取り込み→内部で統合し→新たな状態を生成する」**という共通の論理構造を持つことである。スケールや自律性の深さは異なるが、この連鎖的な情報処理こそが意味生成の核心であり、ホワイトヘッド哲学が生命現象の記述に援用できる理由でもある。
二種類のモデル案:数式と設計の骨格
モデル1:反応拡散系+エージェントベースモデル
化学物質濃度場を偏微分方程式で記述しつつ、個々の細胞を離散エージェントとして扱う混合モデルである。空間を格子状に離散化し、各セルの化学物質濃度を有限差分法で更新する。各エージェント(細胞)は近傍の濃度勾配を計算し、バイアス付きランダムウォークで移動する。
このモデルの特長は、集団レベルのパターン形成(クラスター形成、波状濃度分布など)と個体レベルの行動(軌跡、転向頻度)を同時に観察できる点にある。パラメータとして走化感受度 χ、拡散係数、ステップサイズを変化させることで、応答の閾値効果や頑健性を評価できる。
モデル2:確率過程モデル+情報理論的指標
ラン・アンド・タンブル運動を確率過程として表現し、センサー出力と行動の間の相互情報量 I(S;B) を逐次計算するモデルである。シミュレーションを重ねることで、時間経過とともに I(S;B) が増加して飽和する様子が観察されると期待される。この飽和値が大きいほど、細胞は環境から「意味のある情報」をより多く行動に結びつけていると解釈できる。
パラメータ調整(感受度・ノイズ強度)を通じて走化性能(勾配追跡速度)と情報指標の関係を調べることで、Mattinglyらの理論限界仮説を模型上で再現・検証することも可能になる。
実験的検証への道筋
数理モデルの妥当性を実験的に担保するには、少なくとも以下のアプローチが考えられる。
走化性実験系: マイクロフルイディックデバイスを用いて化学勾配下での大腸菌の運動をトラッキングし、勾配強度と移動速度の関係、ならびに蛍光レポーターによるシグナル伝達活性の時系列計測を行う。これにより I(S;B) に対応する実測指標との比較が可能になる。
自律性・適応実験系: 栄養欠乏などのストレス条件下で単細胞または細胞シートの代謝応答を観察し、代謝マーカーの時間変化や自己維持時間などをモデル予測と照合する。
人工・ロボット系への応用: 人工細胞(再構成代謝回路を持つリポソーム)や光走性ロボット群によって、意味生成の最小系を人工的に構築・検証する試みも有望である。これらは「出来事の連鎖として行動が創発する」というホワイトヘッド的世界観を工学的に実装する可能性を持つ。
まとめ:意味生成の最小モデルが示す視野
本記事で見てきたように、走化性と細胞自律性は単なる生物現象にとどまらず、情報を取り込み・統合し・行動へ変換するという意味生成の最小形として理解できる。そしてこの構造は、ホワイトヘッドが「実際的偶有」として定式化したプロセス哲学の核心と驚くほど対応している。
Keller–Segel方程式による空間パターン形成から、相互情報量 I(S;B) による情報処理効率の定量化まで、複数のモデルを組み合わせることで「意味生成の最小単位」に迫る研究基盤が整いつつある。今後は細胞ネットワークや多細胞系への拡張、さらにはホワイトヘッド的世界観との理論的統合を深めていくことが、生命・認知・哲学を横断する新たな研究地平を開く可能性がある。
コメント