「時間を感じる脳」を光で操作する——なぜ今、時間知覚研究が重要なのか
人間はなぜ、退屈なときに時間が長く感じ、集中しているときに短く感じるのか。この日常的な問いの背後には、脳が「主観的時間」を能動的に生成しているという事実がある。時間知覚の神経基盤を解明することは、パーキンソン病や統合失調症など時間感覚の歪みを伴う神経精神疾患の理解にもつながる、基礎・臨床両面で価値の高いテーマだ。
近年、光遺伝学(オプトジェネティクス) の技術革新により、特定の細胞型を生きた動物でミリ秒精度に操作することが可能になった。これにより「どの回路が時間知覚を担うか」という相関研究から、「その回路を操作すると時間感覚が実際に変わるか」という因果的検証へと研究水準が大きく引き上げられている。
本記事では、現時点で最も再現性が高く因果的証拠が蓄積している三つの回路——中脳SNcドーパミン系、背内側線条体D1/D2-MSN、視床内側背側核(MD)——を軸に、標的選定から実験設計・遺伝子ツール・統計計画まで、研究を実際に立ち上げる際に必要な知識を体系的に整理する。
時間知覚の神経回路——古典モデルから分散システム観へ
皮質–線条体ループ中心モデルの限界
かつて interval timing(秒〜数十秒スケールの時間推定)の神経基盤は、皮質–線条体ループが中心仮説だった。前頭皮質のニューロンが発振活動を生成し、線条体がそれを積算・検出するという「striatal beat frequency モデル」がその代表格だ。このモデルは多くの行動・電気生理データと整合し、今も説明力を持つ。
しかし近年の研究は、時間知覚をより広い分散型ネットワークとして捉えることを支持している。前頭皮質・視床・小脳の相互作用が組み込まれ、単一のペースメーカー仮説では説明しきれない現象が相次いで報告されている。特に注目されるのが、ドーパミン系の因果的役割と視床の精度調整機能だ。
ドーパミンが「時計の速さ」を制御する
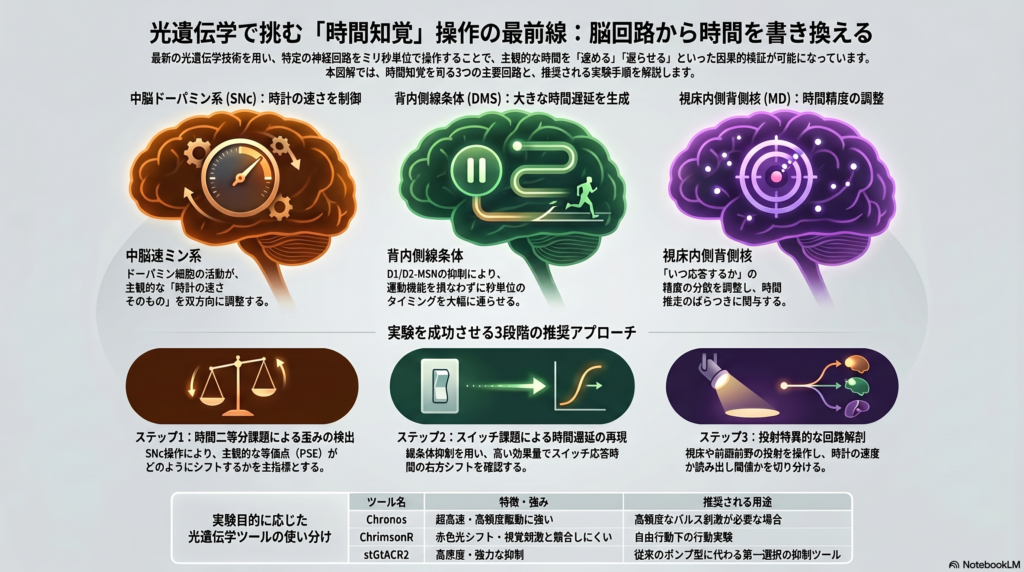
現時点で最も再現性が高く因果推論に向いている回路は、中脳ドーパミン系、背内側線条体、内側前頭前野–視床内側背側核系だ。なかでもSoaresらの研究は決定的で、SNcドーパミンニューロンの一過性活性化・抑制が時間判断を双方向に変えることを示し、ドーパミン活動が時間判断を直接制御しうることを示した。
これはいわば「時計の速さそのものをドーパミンが調整する」という発見であり、従来の「ドーパミンは動機づけや運動制御に関わる」という理解を大きく更新するものだ。ドーパミン活動が主観的時間の進み方を因果的に調整することを示した重要な転換点と位置づけられている。
線条体D1/D2-MSNの役割——効果量の大きな時間遅延
最近のeLife論文では、背内側線条体のD2-MSNとD1-MSNが抑制されるとスイッチ応答時間が遅れることが示された。しかも、鼻先保持時間や移動時間などの課題特異的運動指標を大きく変えずに起こることが示されている。
この「運動を変えずに時間推定だけを遅らせる」という解離が、実験の証拠価値を高めている。効果量は非常に大きく、D2-MSN抑制でCohen’s d=1.7、D1-MSN抑制でd=1.4が報告されている。
視床内側背側核——時間精度の調整器
視床内側背側核の抑制でもピーク時刻の右方シフトと時間幅の増大が生じ、運動そのものではなく時間処理に重要であることが示されている。MD視床は「いつ応答するか」の精度を調整する役割を担う可能性があり、時間推定の分散を調べる課題と特に相性がよい。
標的回路と遺伝子ツールの選定——何を、どう操作するか
優先度別ターゲットマップ
実験の目的に応じて標的を選ぶことが重要だ。標的選定の優先度は、SNcドーパミン系が「主観的時間の進み方そのものを双方向に歪める第一候補」として最高優先度に位置づけられ、背内側線条体D1/D2-MSNが「秒スケールのタイミングを大きな効果量で改変し運動混乱因子が比較的小さい」として同じく高優先度となる。
さらに、内側前頭前野は「timing rule、注意、時間証拠の読み出し」という観点から、投射特異的検証に有望な中優先度ターゲットとして位置づけられており、前頭–視床–線条体の三者連関を回路階層レベルで解剖することも今後の展望として有力だ。
励起ツール——Chronos・ChrimsonR・ChRmineの使い分け
古典的なChR2(H134R)は実績が豊富だが、新世代のツールはそれぞれ特化した強みを持つ。
Chronosは超高速で高頻度駆動に強く、ChrimsonRは赤シフトで視覚刺激との競合を避けたい自由行動実験に向き、ChRmineは非常に大きな光電流と高感度を持ち低光量で大きな効果を得やすい。
ChRmineは深部標的や投射末端の刺激に特に有力だが、過剰駆動とdepolarization blockのリスク管理が必要な点には注意が必要だ。多色実験を計画する場合は、ChrimsonRとChronosを組み合わせて励起と抑制を独立に制御する設計が有効になる。
抑制ツール——stGtACR2が現世代の第一選択
技術面では、抑制にはsoma-targeted GtACR2(stGtACR2)を優先すべきで、従来のhalorhodopsin/Arch系よりも高感度で、軸索側の望ましくない励起を減らした強力な抑制が可能だ。
従来のeNpHR3.0やArchTはポンプ型であるため、長時間照射時に光量・pH・温度の問題が生じやすい。新規実験を立ち上げる場合は、stGtACR2を第一選択にしつつ、先行研究との直接比較が必要な場合にのみhalorhodopsin群を少数併設する設計が合理的だ。
Cre系統と発現系の選択
主力となる遺伝子ツールはCre系統+AAV-DIOが最も実装しやすい。C57BL/6J背景はtiming特性が安定しており、peak-interval procedureに適している。Cre系統としてはDAT-Cre、Drd1a-Cre、Adora2a-Cre/D2-Cre、Vglut2-Creが主候補だ。
プロモーターの選択も慎重に行う必要がある。CaMKIIαプロモーターは厳密な興奮性ニューロン限定ではなく、皮質抑制性細胞にも発現しうるため、細胞型特異性を前提にした解釈には注意が必要だ。hSynプロモーターはニューロン特異性が高く、細胞型操作を目的とした実験での基本選択として推奨される。
行動課題の設計——「時間知覚」と「時間行動制御」を切り分ける
一つの課題ですべてを測ろうとしないこと
時間知覚研究でよく犯されるミスが、一つの行動課題で「知覚」「蓄積」「意思決定」をすべて混在させてしまうことだ。時間知覚は、少なくとも「短いintervalの知覚的分類」と「秒〜数十秒の時間に基づく行動制御」に分けて設計した方が、結論が明確になる。
時間二等分課題——主観的時間の歪みを直接検出する
時間二等分課題は、動物が「短い」か「長い」かを二択で分類する課題であり、**心理測定曲線のPSE(主観的等価点)**のシフトを主指標とする。主計画Aとして、DAT-Creマウスを使いSNcドーパミンニューロンを操作し、時間二等分課題でpsychometric curveのPSEがどう動くかを主指標にすることが、主観的時間の歪みを最短距離で示す方法だ。
スイッチ課題——効果量の大きい時間遅延の再現
主計画Bとして、D1-Cre/Adora2a-Creによる背内側線条体抑制と6–18秒スイッチ課題の組み合わせは、switch response timeのシフトを主指標とし、効果量が大きく、まず確実に「時間を遅らせる」現象を出したい場合に最適だ。
ピークインターバル法——精度と分散を同時に評価する
C57BL/6Jで複数のintervalを用いたpeak-interval procedureは、単なるmotor latencyではなくtiming原理そのものを検証しやすくなる。MD視床の抑制実験など、時間推定の「精度」そのものに介入する仮説を検証する場合に特に向いている。
実験プロトコルの要点——再現性を確保するための実務的注意点
対照群の設計が実験の信頼性を決める
対照群として少なくとも、opsin陰性だが同じCre・同じ光条件の群、蛍光レポーターのみの群、同等のtethering/handlingを受ける対照を含める必要がある。
光そのものが追加の時間手がかりとして機能しないよう、チャンバー内の照明条件を統一し、光のオン・オフが行動上の弁別刺激になることを防ぐ工夫も必要だ。
刺激ウィンドウの細分化——どこで操作するかが仮説を決める
少なくとも、light-off、whole-interval light-on、early-window light-on、mid-window light-on、late-window light-onのtrial typeを前もって固定することにより、clock initiation、accumulation slope、decision thresholdを切り分けやすくなる。
これにより「時計がスタートしないのか」「積算が遅いのか」「読み出し閾値が上がったのか」という仮説をデータで区別できる。
運動共変量の同時記録が「時間知覚操作」の証明を支える
セッションごとに、omission rate、reward count、movement velocity、nosepoke duration、traversal timeを確認することが必須で、DMS研究ではタイミングは遅れたがnosepoke durationやtraversal timeは大きく変化しなかった。ここを再現できないと「時間知覚操作」より「運動撹乱」の可能性が高くなる。
統計計画とサンプルサイズ——混乱因子を数値で切り分ける
一般化線形混合モデルの活用
時間二等分課題のデータは、試行レベルで一般化線形混合モデルを用いることが推奨される。Interval durationを対数変換した説明変数と刺激条件、その交互作用項を含め、マウスを変量効果とすることで、個体差を適切に制御しながらPSEのシフトを推定できる。
within-subject設計を基本とする
paired designで真の効果がdz≈0.8なら80%powerに概ね15匹前後、dz≈0.6なら20匹台前半が必要になる。そのため、本研究ではwithin-subject designを基本にすべきだ。
SNc×二等分課題では14–18匹/群、DMS D1/D2-MSNではそれぞれ12–14匹、MD inhibition×PIでは12–16匹、opsin-negative controlsでは8–10匹が推奨される。新規立ち上げでは発現失敗やmis-targetが現実的に起きるため、理論必要数より多めに設定することが現実的だ。
必須の混乱因子コントロール
混乱因子として特に重要なのは運動・覚醒・報酬期待・感覚cue・光加熱であり、opsin-negative+identical light condition、trial randomization、movement covariateの同時計測、光量の先端実測、実験者の条件盲検が必須だ。
代替ニューロモジュレーションの位置づけ
化学遺伝学(DREADD)——ブロックデザインの補助に
DREADDは秒以下精度が不要なセッション全体への介入に向く。現在はDCZがCNOよりpotent・selective・metabolically stable・fast-actingとされており、chemogenetic block designを使うならDCZを優先すべきだ。ただし試行単位の時間窓操作には使えないため、光遺伝学との補完関係で運用するのが合理的だ。
超音波・磁気刺激——将来展望として
経頭蓋集束超音波(tFUS)はsubmillimeter級の焦点性と深部到達を持つ有望な手法だが、interval timingを直接操作したrodent文献はまだ乏しいため、主役ではなく将来拡張と考えるべきだ。ヒトではrTMSによる時間知覚歪みの報告があるが、マウスではコイル焦点性の問題から主要な手法とは位置づけにくい。
予算と実装コスト——立ち上げの現実的な目安
共有設備がある施設を前提に、光源・rotary joint・patch cable/cannula・operant chamber・AAVを揃えると概ね10,000–18,000 USDで始められる。stereotaxic frameとmicroinjection pumpを自前で新調すると20,000–30,000 USD台が現実的で、これにphotometryを加えると+6,800 USD以上を見込むべきだ。
低コスト化にはopen-source Arduino制御の行動箱やpyPhotometryなどの活用が有効で、完全自作に近い構成を組めばコストを相当圧縮できる。
まとめ——三段階の実験構成が最短距離
光遺伝学による時間知覚の選択的操作研究は、SNcドーパミン系・背内側線条体・MD視床という三つの回路に強い因果証拠が蓄積しており、今がこれらを統合的に検証する好機だ。
実験を立ち上げる際の推奨順序は明確だ。まずDAT-Cre×SNc操作×時間二等分課題でPSEシフトを示し、次にD1-Cre/D2-Cre×背内側線条体抑制×スイッチ課題でswitch timeの右方シフトを再現し、最後にMD視床またはPFC→DMS/PFC→MD投射特異的操作で「時計速度」か「蓄積・読み出し」かを回路レベルで切り分ける。この三段構えが、既存文献の最も強い部分を再現しつつ現状のギャップに答える設計として最も堅実だ。
対照群・運動共変量・光量管理・組織学検証を丁寧に積み重ねることで、「時間を変えた」という主張に対する反論を先回りして封じることができる。それが、この研究テーマで信頼される論文を書くための最短ルートでもある。
コメント