はじめに:発生プログラムの変化が生み出す形態進化
進化発生生物学の中心的な問いは、「遺伝子の変化がどのようにして生物の形態を変えるのか」という点にある。特に脊椎動物では、ホメオティック遺伝子であるHox遺伝子群が胚の前後軸に沿った体制を決定する鍵を握っている。興味深いのは、Hox遺伝子そのもののタンパク質配列は種を超えて高度に保存されているにもかかわらず、魚類のヒレから四肢動物の手足への進化、ヘビの四肢消失、トゲウオの骨盤退化など、劇的な形態変化が実現している点である。
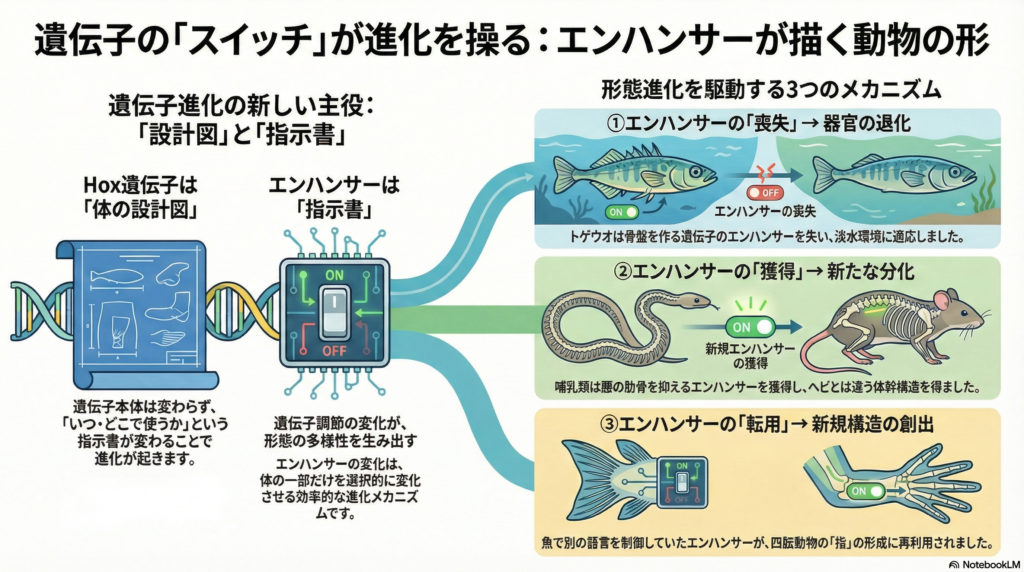
こうした形態の多様化を可能にしたのが、Hox遺伝子の発現を制御するエンハンサー領域の進化的再構成である。エンハンサーは遺伝子からしばしば数万塩基離れた位置に存在するシス調節配列であり、その変異や欠失、新規獲得によって遺伝子の発現パターンが変化する。本記事では、Hox遺伝子とそのエンハンサーの進化が脊椎動物の形態形成に及ぼした影響について、魚類から四肢動物まで具体的事例を交えながら解説する。
Hox遺伝子の基本構造とコリニアリティの原理
Hox遺伝子クラスターと時空間的発現制御
Hox遺伝子は染色体上にクラスターとして配置され、その物理的配置と発現領域に対応関係が認められる。これをコリニアリティと呼び、クラスター上で3’側に位置する遺伝子ほど胚の前方で早期に発現し、5’側の遺伝子ほど後方で後期に発現する傾向がある。この時空間的な発現制御は体節形成と密接にリンクし、各Hox遺伝子の発現境界位置によって体節や器官のアイデンティティが決定される。
脊椎動物では全ゲノム重複を経てHoxクラスターが4セット(HoxA~HoxD)に増加し、哺乳類では39個のHox遺伝子を持つ。硬骨魚類ではさらなる重複を経験し、ゼブラフィッシュでは7クラスターを保有する。重複により生じた同系統の遺伝子(パラログ)は機能的冗長性を持つため、単一遺伝子の喪失では表現型が顕在化しにくい。しかし全パラログ群の機能を同時に失うと、顕著なホメオティック変換が現れる。
全ゲノム重複がもたらした機能的冗長性と進化可能性
マウスでHox10群の3遺伝子全てを不活性化すると、本来肋骨を持たない腰椎・仙椎領域に肋骨が伸長する異常が生じる。これはHox10パラログが正常発生では腰域で肋骨形成を抑制していることを示し、種間差においてもHox発現境界の変化が脊椎の肋骨パターンを変えうることを示唆する。このように、遺伝子重複による冗長性は一見すると進化的制約に見えるが、実際には一部のパラログが新機能を獲得する余地を与え、形態進化の可塑性を高める役割を果たしている可能性がある。
エンハンサー領域の再構成が形態進化を駆動するメカニズム
エンハンサー新規獲得による発現領域の拡大
エンハンサーの新規獲得は、遺伝子が従来発現していなかった組織で活性化される契機となる。哺乳類では腰椎領域でHox10遺伝子群が強く発現し肋骨形成を抑制しているが、これは他の爬虫類と比べ腰部特異的なHox発現制御エンハンサーを獲得した結果と考えられる。ヘビでは体幹軸全体に肋骨が存在するが、哺乳類や鳥類では胸郭より後方の腰部で肋骨が消失する。
古典的にはヘビでHox遺伝子群の機能喪失が起きたため体軸の分節パターンが単純化したと考えられていたが、実際には哺乳類や鳥類がHox発現の新たな調節を獲得することで、頸・胸・腰など明瞭な領域分化を進化させたとする報告がある。これは新規エンハンサー獲得による既存遺伝子の発現領域追加が形態の複雑化を導いた事例である。
エンハンサー喪失による器官退化のメカニズム
既存のエンハンサー配列が変異蓄積や欠失によって機能低下すると、遺伝子の発現が限定され対応する構造が退化する。トゲウオの複数の淡水集団では骨盤ヒレが縮小・消失する適応進化が起きているが、その原因はPitx1遺伝子の骨盤部位エンハンサーが繰り返し欠失突然変異により破壊されたためである。Pitx1自体のタンパク質配列には変化がないにもかかわらず、該当エンハンサーの欠失で発現が局所的に消失し、それに伴い骨盤構造が退化した。
Pitx1のケースでは胚の他組織では発現が維持され致死的影響なく形質だけが変化するため、適応進化に有利な遺伝的変化となった。同様にヘビ類の四肢消失に際しては、四肢芽でのSonic hedgehog遺伝子発現を駆動する遠隔エンハンサー(ZRS)に塩基置換の蓄積・部位欠損が生じ、エンハンサー活性が機能喪失したことが原因の一つである。
エンハンサー転用による新規構造の創出
もともと異なる文脈で機能していたエンハンサーが、新たな遺伝子や組織の制御に流用される現象をエンハンサー転用と呼ぶ。四肢動物の指趾形成に必要なHoxdクラスター遠隔エンハンサー群は、元来は脊椎動物の総排泄腔付近の発生制御に働いていたものが四肢進化の際に新たに末端骨形成に転用されたという仮説が近年示された。
ゼブラフィッシュにおいて該当するHoxdクラスター下流域を全域欠失させる実験では、ヒレの遠位部形成には影響がない一方で総排泄腔の形態異常が引き起こされた。つまり魚類ではこのエンハンサー領域は本来総排泄腔に隣接する尿生殖器官形成を制御していたが、四肢動物では同じ領域が指骨形成に必要な調節ドメインとして再利用されたと考えられる。このようなエンハンサー転用は、既存の発生ネットワークを新しい構造の創出に活用する効率的な進化メカニズムを示している。
魚類のヒレから四肢動物の手足への進化プロセス
深い相同性とHox13遺伝子の役割
四肢動物の四肢骨格は約3億6千万年前のデボン紀に魚類のヒレから進化したとされる。マウスでHoxa13とHoxd13の両方を欠失させると手足の指骨が完全に欠如することから、これらHox13遺伝子は四肢の末端構造形成に必須である。同様のHox13遺伝子群をゼブラフィッシュで同時変異させた実験では、ヒレの先端部分が欠失することが報告され、魚類にも四肢動物の指に対応する遠位構造形成の遺伝子ネットワークが潜在することが示された。
この現象は深い相同性の一例であり、四肢動物の新奇形質である指趾が、実は祖先魚類から受け継がれた発生プログラムを再活性化・拡張したものである可能性を示す。しかしながら、魚類のヒレには明瞭な指骨は存在せず、その遠位骨格は鰭条と呼ばれる真皮性の支持要素に留まる。
HoxDクラスター調節領域の進化的転用
四肢動物ではHoxDクラスターの5’側調節領域が指形成に不可欠であり、これを削除すると指骨は全く形成されない。ゼブラフィッシュのHoxDaクラスターに隣接する調節領域を欠失させたところ、魚ではヒレの形態異常は起きず、代わりに顕著に現れたのは総排泄腔の形成不全であった。このことは、魚類ではHoxクラスター下流域のエンハンサー群が本来ヒレではなく総排泄腔の発生制御に使われていたことを示唆している。
研究者らは「四肢動物の指形成に利用されている調節ランドスケープは、元来は祖先脊椎動物の総排泄腔に固有の調節モジュールであり、それが四肢進化時に丸ごと転用された」との仮説を提唱した。この転用により、四肢動物では指骨形成に必要な遺伝子群の発現がヒレよりも拡張され、明確な指の形成が可能になったと考えられる。
トゲウオの骨盤ヒレ退化に見るエンハンサー欠失の進化的意義
Pitx1遺伝子エンハンサーの機能喪失
海産の祖先型イトヨは発達した骨盤を持つが、淡水湖に複数回独立に侵入した集団では骨盤が小型化あるいは完全に消失する進化が起こった。遺伝解析により骨盤サイズの変異はPitx1遺伝子を含む座位に強く支配されることがわかった。興味深いことに、骨盤退化型のイトヨではPitx1遺伝子のタンパク質配列には変化がなく、胚発生におけるPitx1の発現そのものが骨盤領域で消失していた。
骨盤欠如型イトヨでは、Pitx1遺伝子の上流約2.5キロ塩基の領域に、海産イトヨと比べ約502塩基対の欠失が見つかった。この部分は骨盤芽でのPitx1発現を担うエンハンサーと考えられ、Pel領域と命名された。
トランスジェニックレスキュー実験による因果関係の証明
研究チームは骨盤退化型イトヨの受精卵に、海産型からクローンしたPelエンハンサーとPitx1遺伝子ミニコンストラクトを導入した。その結果、導入個体では腹側に本来消失していたトゲ(骨盤棘)が再形成された。骨染色標本では、導入魚では腰帯に前後の枝や棘を伴う大型の骨盤骨格が再建されているのに対し、非導入の元集団では痕跡的な小片しか形成されていない。
このトランスジェニックレスキュー実験は、失われたPitx1エンハンサーさえ復元すれば骨盤構造を蘇らせ得ることを示し、エンハンサー喪失が形態変異の直接原因であることを強力に裏付けた。イトヨの例は自然選択下でエンハンサーの欠失が反復的に起こり、同一の形態収斂をもたらした顕著な事例である。
ヘビの体制進化とHox遺伝子発現パターンの再解釈
古典的仮説の見直し:Hox機能喪失説からの転換
ヘビ類は四肢の消失と極端な胴体の伸長というユニークな体制を進化させた。古典的には「ヘビの体はHox遺伝子制御の破綻によって領域分化が失われた結果」だと推測されてきた。しかし詳細な形態解析により、ヘビにも他の有鱗目と同じく複数の椎骨レギオン(頸部・前胴部・後胴部・尾部など)が区別できることが明らかになった。
各領域の境界はニシキヘビのHox遺伝子発現パターンでマッピングされた位置と対応しており、ヘビでもHoxコードが通常通り機能して胴体をパターン形成していることが示唆された。むしろ哺乳類や主竜類の方が、頸部や腰部などの明瞭な分節パターンを進化的に獲得した特殊例であり、ヘビ類は基本的なパターンに留まっている可能性が高い。
Sonic hedgehogエンハンサーの段階的劣化
ヘビで四肢が消失した直接の遺伝学的原因として、Shh遺伝子エンハンサー(ZRS)の退化が主要因とみられる。基盤的なヘビ(ボアやニシキヘビ)ではZRSが他の脊椎動物同様に保守的であるのに対し、進化的に派生したヘビ(コブラやガラガラヘビなど)ではZRS塩基配列の突然変異蓄積が顕著である。
マウス胚にこれら各種のZRS配列を組み込んだトランスジェニック実験では、原始的ヘビのZRSは後肢芽でわずかな活性を残すのに対し、派生的ヘビのZRSは全くShh発現を誘導できないことが示された。わずか1箇所、ヘビ系統で失われた転写因子結合サイトを元に戻すだけでエンハンサー活性が蘇り正常な肢が形成されたという逆実験の結果もある。
コリニアリティ仮説から発現のホメオーシスへのパラダイムシフト
古典的コリニアリティ仮説の限界
古典的コリニアリティ仮説では、Hoxクラスターの構造と発現順序が強く進化的拘束を受けるため、胚の基本的な体節配置は容易には変化しないと考えられていた。脊椎動物の胚は発生中盤の定型発生期において類似した体制を示し、この時期にHox遺伝子が前後軸に沿って順次発現して基本的な領域区画が形成される。進化的多様化は、その定型段階以降の発生過程でHox遺伝子の発現タイミングや持続期間が変化するヘテロクロニーによって実現すると提唱された。
空間的コリニアリティの変化と形態進化
その後の分子発生学的研究は、空間的コリニアリティの変化、すなわちHox遺伝子の発現領域そのものの前後転移や拡大・縮小もまた形態進化に寄与しうることを示している。ヘビの胴体パターンや鳥類の頸椎数変化などはHox発現領域の移動(シフト)として理解でき、これはまさに「発現領域のホメオティックな転換」と言える。
Hox遺伝子それ自体の配列や相互作用ネットワークは保存されつつ、そのスイッチの入れ方(いつ・どこで発現するか)が進化で調節され直すことで多様な体制が生み出された。この理解は、コーディング変異よりシス調節変異が重要という「進化における遺伝子調節パラダイム」の一端を成すものであり、古典的モデルを修正・包括する新たな理論枠組みとして定着しつつある。
CRISPR-Cas9技術がもたらす進化発生学の新展開
非モデル生物での機能検証が可能に
エンハンサー領域の再構成と形態進化の因果関係は、近年のCRISPR-Cas9技術の飛躍的発展によって直接検証できる時代に入った。従来、発生段階での遺伝子機能を調べるには主にノックアウトマウスなどモデル生物の作製が必要であったが、CRISPR技術により非モデル生物でも特定配列の変異導入が可能となり、進化的変異の再現実験が飛躍的に容易になっている。
ゼブラフィッシュHoxエンハンサー欠失実験、イトヨPitx1エンハンサー復元実験、マウスにおけるヘビ型ZRSエンハンサー機能検証は、いずれもゲノム編集・トランスジェニック手法を組み合わせることで進化上の仮説を直接試すことに成功した例である。
進化を巻き戻す実験の実現
CRISPRを応用したマウス胚へのエンハンサー置換というアプローチで、ヘビのZRS配列をマウスゲノム上の対応位置にノックインし、そのマウスが四肢欠損になることを示した研究がある。さらにそのマウスの変異エンハンサー配列中の1塩基を先祖型に戻す操作も行い、失われた後肢が正常に再生することを確認している。このような「進化を巻き戻す」実験は、ゲノム編集技術があって初めて成し遂げられたものであり、進化発生学に新次元の証拠をもたらした。
CRISPRは多遺伝子同時変異を可能にする点でも革新的である。Hox遺伝子は機能冗長性が高く単一変異では効果が見えにくいが、ゼブラフィッシュのHoxクラスター7箇所すべてに変異を導入し、初めて魚類で明瞭なホメオティック変異体を作出することに成功した例もある。
まとめ:形態進化におけるシス調節変異の中心的役割
脊椎動物の形態進化において、Hox遺伝子そのもののタンパク質配列は高度に保存されている一方で、その発現を制御するエンハンサー領域の変化が劇的な形態変化を可能にしてきた。エンハンサーの新規獲得は発現領域の拡大と新規構造の創出を、エンハンサー喪失は器官退化を、エンハンサー転用は既存の発生プログラムを新しい文脈で活用することを可能にした。
魚類のヒレから四肢動物の手足への進化、トゲウオの骨盤ヒレ退化、ヘビの四肢消失といった具体的事例は、いずれもエンハンサー領域の再構成が形態進化の主要な駆動力であることを示している。古典的なコリニアリティ仮説が強調したHoxクラスター構造の厳密な保存性に対し、現代のEvo-Devoはむしろ発現パターンの可塑性とシス調節エレメントの進化可能性を重視する。
CRISPR-Cas9などのゲノム編集技術の発展により、進化上の推定シナリオを生きた胚で再現・検証する試みが現実のものとなった。今後、特定の種で失われた器官をエンハンサー改変で復元したり、新たに進化した形質を逆に他種に導入したりといった実験がさらに活発化すると期待される。こうした「進化の実験再現」は、生物の形態進化における遺伝的メカニズムを厳密に突き止める上で極めて有力な手段であり、Evo-Devo研究を理論から実証の段階へと深化させている。
コメント