量子生物学とは何か——生命現象に潜む量子効果の実像
「量子」という言葉はしばしば神秘的なニュアンスで語られがちですが、量子生物学はあくまで厳密な実験科学です。光合成のエネルギー移動や鳥の方位感覚を支える分子機構が、古典物理だけでは説明しにくい量子力学的ダイナミクスを活用している可能性が、超高速分光やスピン化学の手法によって示されつつあります。一方で、「量子効果が意識を生み出す」という主張は、実証レベルでは大きな開きがあります。本記事では、比較的証拠の整った光合成・鳥類磁気感知の知見を「参照点」に、脳・意識研究における量子仮説の現在地を批判的に整理します。
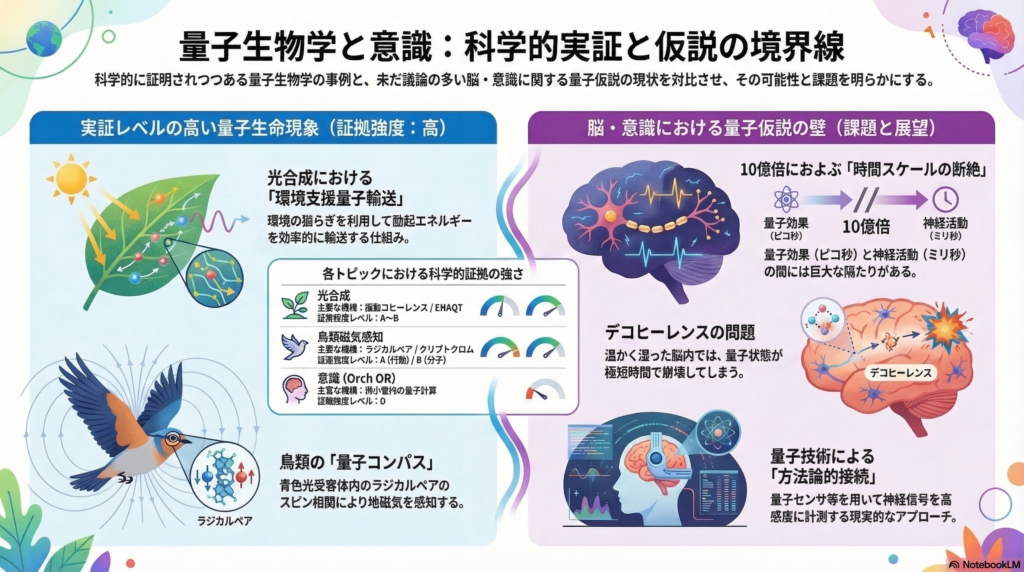
光合成における量子コヒーレンス——2次元電子分光が示すもの
ピコ秒以下の世界で何が起きているか
光合成の光捕集過程では、光子を吸収して生じた励起(エキシトン)が、色素—タンパク質複合体を伝って反応中心へ到達し、最終的に化学エネルギーへ変換されます。この励起エネルギー移動(EET)は、フェムト秒からピコ秒という極めて短い時間で進行します。
2000年代以降、2次元電子分光(2DES)という超高速分光技術の発展により、緑色硫黄細菌が持つFMO複合体などで「コヒーレンス様のビート信号」が観測されました。当初、これは「量子的な波束がタンパク質内を伝わっている証拠」として注目を集め、「光合成は量子コンピュータのように動いている」という表現がメディアに広まりました。
「電子コヒーレンス」か「振動コヒーレンス」か——解釈論争の現在地
しかしその後の研究により、解釈はより慎重になっています。2DESで観測される長寿命の振動成分の多くは、純粋な電子コヒーレンスではなく、電子と分子振動が混ざり合った「ビブロニック(電子—振動混成)」状態、あるいは純粋な振動起源である可能性が指摘されています。室温での電子デコヒーレンスは数十フェムト秒程度に急速に減衰するとする再解析も存在し、「室温で長寿命の純粋電子コヒーレンスが生体機能に必須」という強い主張は、証拠強度としてはB〜Cレベルに落ち着きつつあります。
より現代的な観点では、「自然は散逸を避けるのではなく、環境揺らぎ(フォノン)との結合を利用して効率的なエネルギーフローを設計している」という開放量子系の視点が中心になっています。適度なデコヒーレンスが輸送効率を高め得るという「環境支援量子輸送(ENAQT)」の理論提案も、この文脈で重要な役割を果たします。
光合成量子生物学の証拠レベルまとめ
超高速分光によってコヒーレンス様シグナルが観測されること自体は再現性が高く、観測事実としての証拠強度は高いと言えます(Aレベル)。一方で、「その量子的ダイナミクスが生物学的機能に”必須”か」という問いへの答えは、系・条件・解析モデルに依存し、依然として議論の余地が残っています。
鳥類の磁気感知——ラジカルペア機構の強力な証拠
行動実験が示す「量子コンパス」の特徴
渡り鳥が地磁気を感知して方位を認識することは古くから知られていますが、そのメカニズムは長年の謎でした。鳥類の磁気コンパスには次のような特徴があります。
- 光依存性がある(暗闇では機能しにくい)
- 磁力線の「極性」ではなく「傾き」を使う傾斜コンパスである
- 地磁気程度の非常に弱い磁場で機能する
これらの性質は、光によって生成されるラジカルペアのスピン化学で説明できます。ラジカルペア機構とは、光照射によって生じた二つの不対電子(ラジカル)の間のスピン相関が、外部磁場によってわずかに変化し、反応生成物の比率が変わることで方位情報が化学信号に変換される、という仕組みです。
とくに決定的な証拠とされるのが、特定の周波数帯(MHz帯)の無線周波数(RF)磁場を照射すると鳥の方位行動が乱れるという実験です。これはラジカルペアのスピン遷移に共鳴的に干渉し得る周波数に対応しており、ラジカルペア仮説と強く整合します。さらに、都市環境の電磁ノイズで方位感覚が失われ、電磁シールドすると回復するという報告は、この機構の生態学的妥当性も支持します。
クリプトクロムCRY4——分子候補の特定が進む
ラジカルペア機構の分子的担い手として、網膜に存在する青色光受容体「クリプトクロム(CRY)」が有力候補として浮上しています。とくに渡り鳥(ヨーロッパコマドリ)に由来するCRY4は、試験管内(in vitro)の実験で地磁気相当の磁場強度に対する感受性を示し、非渡り種(ニワトリ、ハト)のCRY4より感受性が高いという報告があります。変異解析では、フラビン—トリプトファン連鎖による複数段階のラジカルペアが磁場効果の生成とシグナルの安定化に関与し得ることも示されています。
「量子効果→化学収率の差→細胞シグナル→神経回路→行動」という因果の鎖が、他のどの量子生物学トピックよりも具体的に描けつつある点が、鳥類磁気感知研究の最大の強みです。ただし、分子レベルの証拠(in vitro)から生体内での知覚(in vivo)への接続、すなわちどの細胞でどの下流シグナル経路を介して視覚系へ情報が伝わるかは、現在も研究が進行中です。
脳・意識における量子仮説の現在地
マイクロチューブリン量子計算仮説(Orch OR)の論点
意識に量子力学が本質的に関わるという立場の代表例が、ロジャー・ペンローズとスチュアート・ハメロフが提唱した「調整された客観的還元(Orch OR)」仮説です。この仮説は、神経細胞内の微小管(マイクロチューブリン)が量子計算の基盤として機能し、その「量子的崩壊」が意識の離散的な瞬間を生み出す、という主張を含みます。
しかし主要な批判は二点に集約されます。第一に、脳内の温度・湿度・イオン濃度という環境条件では、量子計算に必要なコヒーレンスが神経ダイナミクスの時間スケール(ミリ秒〜秒)まで保たれるとは考えにくく、デコヒーレンスが極短時間で起きるとする理論的見積もりが多数提示されています。第二に、神経回路レベルで説明可能な現象に対する「量子を必要とする独自予測」が弱く、実験的に反証可能な形の提案が限られています。
現時点で「微小管で神経生理学的時間スケールの量子コヒーレンスを実測し、意識変数と因果的に結びつけた」強いデータは乏しく、証拠強度はDレベルにとどまります。
核スピン量子情報仮説——長寿命コヒーレンスの可能性と課題
電子スピンよりも環境との結合が弱い核スピンは、理論的には長寿命のコヒーレンスを保ちやすいと考えられます。この観点から、リン酸イオンや「Posner分子」(Ca₉(PO₄)₆)を脳内の量子メモリとして位置づける理論提案があります。
概念としての面白さはあるものの、「脳内でこれらの分子が実際に生成・安定化・輸送・読み出しされるか」という問いに答える実験的証拠は現状では不十分です。化学的実在性や構造に関する反証的論点も含め、現象の確立にはほど遠い状況です(C〜Dレベル)。
キセノン同位体と麻酔——スピン仮説への間接的示唆
より実験的に具体性がある話題として、キセノン(Xe)の核スピンを持つ同位体と持たない同位体で麻酔効力が異なる可能性を示した動物実験があります。外電子殻が同一の同位体間で差が観測されたとすれば、物理的に興味深い現象です。
ただしこの結果は、独立した追試が不足していること、麻酔の作用点は多標的(受容体・膜・タンパク質疎水空洞など)であること、スピン差が「どの分子反応」を経て意識変数へ影響するかが未同定であること、という理由から、現時点では証拠強度はCレベルとされます。
量子効果と神経生理学——時間・スケールの隔たりという根本問題
量子生物学と意識研究をつなぐ最大の障壁は、時間スケールの圧倒的な差です。
光合成のエネルギー移動はフェムト秒〜ピコ秒で完結します。鳥類磁気感知のラジカルペア反応はマイクロ秒以上の時間窓を必要とします。一方で、神経の活動電位は約1ミリ秒、シナプス伝達の遅延は0.5〜1ミリ秒、意識の神経学的指標(大域的統合、ガンマ振動など)はミリ秒〜秒のスケールで動きます。
光合成の量子的時間窓と意識の神経学的指標の間には、10億倍以上のスケールの差があります。これは「量子が意識を直接作る」という考え方に対する根本的な制約です。
より現実的な「接続の型」は、鳥類磁気感知が示すように、「量子効果→化学収率の差→細胞シグナル→神経回路→行動・知覚」という増幅の連鎖を脳内で具体的に同定することです。この鎖が成立しない限り、量子コヒーレンスがたとえ分子レベルで存在したとしても、神経機能に現れない可能性が高い、というのがデコヒーレンス論の核心的指摘です。
意識理論との接続——GNWとIITは量子を必要とするか
主流の意識理論は古典的神経回路で成立する
グローバル・ニューロナル・ワークスペース(GNW)理論は、再帰的な神経回路処理と「イグニッション」(情報が大域的に利用可能になる過程)が、報告可能な意識経験の神経基盤であるという枠組みです。統合情報理論(IIT)は、経験の公理から「因果的統合度(Φ)」という指標を導き出し、意識の程度と同一視しようとします。
これらはどちらも基本的に古典的な神経回路ダイナミクスで成立し、量子的自由度を原理的に必要としません。したがって「量子機構が意識に本質的に関与する」と主張するためには、少なくとも「主流の古典的理論が説明しきれない現象を量子機構がより精確に予測できる」という証拠が必要になります。
量子を組み込む三つの現実的なアプローチ
現時点で比較的妥当とされる接続の形は三つあります。
**①実装中立型(弱い接続):**意識理論(GNW/IIT)は古典的ですが、その基盤となる分子が量子的制約を受け、結果として回路ダイナミクスに影響する。鳥類磁気感知はこの型の現実例であり、量子効果は感覚入力の「前段」で機能します。
**②量子資源必須型(強い接続):**コヒーレンスやエンタングルメントが意識計算の本質的操作に必要という立場(Orch ORなど)。強い反証可能性を持つはずですが、実験的アクセスが難しく、現状では証拠が薄いです。
**③量子計測・技術主導型(方法論的接続):**量子センサや超偏極MRI/NMRで神経系の微視的シグナルを高感度に計測し、主流意識理論の検証力を上げる。日本の量子生命科学政策が推進する方向性と一致しており、最も現実的なアプローチです。
実験的に検証可能な予測と今後の研究戦略
「量子が意識に関与するか」を適切に検証するには、量子自由度の同定と神経・行動への因果接続を切り分けることが必要です。現時点で実現可能性と決定力のバランスが取れていると考えられる実験方向は以下の通りです。
**スピン化学仮説の検証:**キセノン同位体麻酔効果の複数施設・盲検による独立追試、特定周波数のRF磁場との組み合わせによる共鳴的依存性の確認、光条件(フラビン系)との相互作用検証。これらは鳥類研究で確立された実験デザインを参照できます。
**CRY4系の定量的検証:**膜結合・分子配向を制御した状態でのin vitro磁場効果の精密測定は、鳥類磁気感知の因果鎖の弱点(分子配向が方位情報をどう保持するか)を直接攻めるアプローチです。
**量子センサの神経系応用:**ダイヤモンドNVセンターなどの量子センサを使った神経系分子シグナルの局所磁場・代謝計測は、意識の直接説明ではなく「意識研究の測定論を前進させる」という意味で高いインパクトが期待できます。
まとめ——量子生物学と意識研究の現在地と今後の方向性
光合成と鳥類磁気感知は、「生体が量子現象を利用し得る」ことを示す強力な事例です。しかしその量子効果は、局所的な分子反応の中で化学収率という「古典的な量」へ写像され、最終的に生理機能へ増幅されるという点に本質があります。
意識研究に量子生物学の知見を接続するうえで現実的な戦略は、「量子が意識の中核構成要素である」という強い主張よりも、量子効果が「感覚入力・調節・ノイズ源」として神経回路へ影響する具体的経路(スピン化学、酸化還元、光受容体、麻酔作用点)を絞り込み、主流の意識理論や麻酔神経科学の実験系で検証することです。
日本の量子生命科学における量子センサ・超偏極NMR・高速2DESといった技術基盤の整備は、「分子イベントを測って因果推論可能にする」という点でこの戦略と完全に整合しており、今後の展開が期待されます。
コメント