はじめに:なぜ野生植物の光周性研究が重要なのか
植物が季節の変化を正確に予測し、適切なタイミングで開花や休眠を開始する能力は、生存と繁殖成功に直結する重要な形質です。特に野生環境では、わずかなタイミングのずれが致命的な結果をもたらす可能性があります。
植物はこの季節予測において、日長という信頼性の高い環境シグナルを利用しています。温度や降水量と異なり、日長は緯度と季節によって規則的に変化するため、植物は光受容体と概日時計を進化させることで、この情報を精密に読み取る仕組みを獲得してきました。
本記事では、野生植物における光周性の分子基盤、概日時計との統合機構、そして生態学的影響について、最新の研究知見を交えながら包括的に解説します。
光周性を支える光受容体システム
被子植物に存在する5種類の光受容体
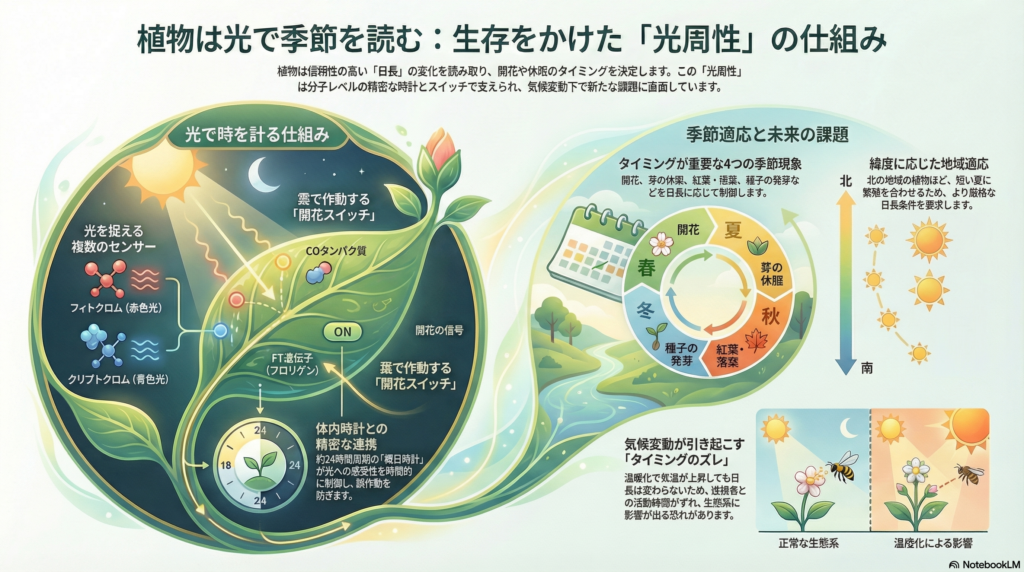
被子植物は光環境を多角的に捉えるため、複数の光受容体システムを進化させてきました。主要な光受容体として以下の5種類が知られています。
フィトクロムは赤色光と遠赤色光を感知し、光周性反応の中心的役割を担います。このタンパク質は光によって活性型(Pfr)と不活性型(Pr)の間を可逆的に変換し、暗期の長さを測定する分子時計として機能します。
クリプトクロムは青色光とUV-A光を検出し、特にシロイヌナズナのCRY2は光周性開花において重要な役割を果たします。その他、フォトトロピン、ZTL/LKPファミリー、UVR8といった光受容体が、それぞれ異なる波長域を担当し、植物の光環境認識を多層的に支えています。
フィトクロムの二重機能:砂時計と光スイッチ
フィトクロムは光周性において二つの異なる機能を持ちます。一つは「砂時計的タイマー」としての役割です。暗期において活性型(Pfr)は徐々に不活性型(Pr)へと戻るため、長い夜が続くとPfrが完全に消失します。短日植物ではこのPfr消失が開花抑制の解除信号となります。
逆に、夜明け時にPfrが残存していると、短日植物では開花が阻害されます。これにより植物は夜の長さを正確に測定し、季節の進行を判断できるのです。この仕組みは、化学反応の進行速度を利用した生物時計の巧妙な例といえます。
CO-FTモジュール:光周性制御の中核メカニズム
葉で作動する分子スイッチ
多くの植物種に共通する光周性の中核機構は、葉で機能するCONSTANS-FLOWERING LOCUS T(CO-FT)モジュールです。このシステムでは、CO転写因子が概日時計によって発現リズムを制御され、誘導的な日長条件下で適切なタイミングで安定化します。
長日植物であるシロイヌナズナでは、COタンパク質が夕方に光と同時に存在するときに安定化し、FLOWERING LOCUS T(FT)遺伝子を活性化します。FTは「フロリゲン」とも呼ばれる移動性シグナルで、葉から茎頂へと輸送され、直接的に花芽形成を誘導する鍵分子です。
クリプトクロムによるCO安定化
クリプトクロム、特にシロイヌナズナのCRY2は、青色光を感知してCOタンパク質を安定化させることでFT発現を促進します。この仕組みは「外的一致モデル」として理解されており、光シグナルが概日時計によって規定された許容時間帯と一致したときにのみ、開花反応が引き起こされるという原理を示しています。
興味深いことに、同じ分子機構が長日植物と短日植物で逆の機能を果たす場合があります。イネやダイズなどの短日植物では、同様の遺伝子が異なる制御ロジックの下で働き、長い夜が開花を誘導するように機能しています。
概日時計との統合:24時間周期が生む精密制御
時計遺伝子が規定する光感受性
光周性の基盤には、植物の内在的な概日時計が存在します。この約24時間周期の生物時計は、光受容体経路と遺伝子発現を時間的にゲート制御しています。
シロイヌナズナでは、ELF3やGIGANTEA(GI)といった時計遺伝子が損なわれると、日長に対する感受性が失われ、植物は日長に関係なく開花します。これはCO遺伝子の発現リズムが破綻し、光周性シグナルを正しく解釈できなくなるためです。
時計周期の変異が生む地域適応
概日時計の周期そのものの長さも、光周性反応に影響を与えます。野生のウキクサ(Lemna)を用いた研究では、概日時計の自由走行周期が長い系統ほど、開花に必要な臨界夜長が長いことが示されています。
この知見は重要な意味を持ちます。すなわち、概日時計の速度そのものが地域適応の基盤となりうるということです。異なる緯度や気候帯に分布する集団では、時計遺伝子の微細な変異が蓄積することで、各地域の日長パターンに最適化された光周性反応を進化させている可能性があります。
光周性が制御する主要な季節現象
開花タイミングの多様な制御様式
光周性は開花タイミングの制御において最も顕著な役割を果たします。植物は大きく長日植物と短日植物に分類されます。
長日植物は夜が短くなると開花します。シロイヌナズナはその代表例で、春から初夏にかけての日長延長を感知して開花を開始します。一方、短日植物は夜が一定以上長くなると開花し、イネやダイズがこのカテゴリーに属します。これらは秋の日長短縮を感知して繁殖段階へ移行します。
同じFTタンパク質が両方のグループで開花促進因子として機能しますが、その上流の制御ロジックが異なることで、正反対の日長応答を実現しています。
芽の休眠と成長停止
落葉性樹木では、短日条件が秋の成長停止と冬芽形成を誘導します。この反応は、来るべき冬季の低温や凍結から植物体を保護するための適応戦略です。
ポプラ属(Populus)を用いた研究では、短日条件下で夜間のLHY2発現が上昇し、FT2遺伝子を抑制することでシュート伸長が停止し、冬芽が形成されることが明らかになっています。この仕組みは草本植物の開花制御と類似した分子基盤を持ちながら、樹木特有の多年生戦略に応用されています。
葉の老化と落葉の制御
光周性は葉の老化、すなわち紅葉や落葉の主要な制御因子でもあります。温度と比較して、日長は冬接近のより信頼性の高い指標とされています。
短日条件下では、葉からの栄養回収プロセスが促進され、その後に離層が形成されて落葉が起こります。この精密なタイミング制御により、植物は霜害を回避しながら、葉に蓄えられた窒素やリンなどの貴重な資源を効率的に回収できます。
発芽における光周性
一部の野生植物では、種子の発芽タイミングも日長によって制御されます。特に乾燥地に生育する植物では、光周性が降雨季節の予測に寄与している可能性があります。
発芽段階での光周性制御は、発芽後の実生が適切な成長期間を確保できるタイミングを見極めるための戦略と考えられます。
光周性形質における地域適応と進化
緯度に沿ったクライナル変異
光周性形質は緯度に沿った連続的な変異、すなわちクラインを示すことが多くの種で報告されています。これは日長パターンが緯度によって系統的に変化するため、各地域の集団が局所的な日長環境に適応した結果です。
日本各地から収集されたLemna aequinoctialisの研究では、北方の系統ほど短日条件を強く要求することが示されています。これは高緯度地域では夏季の日長が極端に長くなるため、より厳格な短日要求性を持つことで、適切な時期にのみ繁殖段階へ移行する適応と解釈されます。
作物化における光周性遺伝子の改変
農作物の祖先である野生植物でも同様の適応パターンが見られます。オオムギやイネでは、光周性遺伝子の機能喪失変異が高緯度地域への栽培拡大を可能にしました。
野生型では厳格な日長要求性により栽培可能地域が限定されますが、光周性鈍感変異体では幅広い緯度で栽培が可能になります。この人為選択の歴史は、光周性が植物の地理的分布を規定する強力な制約であることを示しています。
気候変動下での光周性:新たな生態学的課題
光的障壁という概念
気候変動により温度帯は地球規模で移動していますが、日長は緯度に完全に固定されています。この根本的な非対称性が、植物の分布変化に新たな制約をもたらす可能性が指摘されています。
植物が温暖化に応じて高緯度へ分布を拡大すると、これまで経験したことのない日長パターンに直面します。光周性シグナルと実際の環境条件がずれる「光的障壁」が生じ、適切なタイミングでの開花や休眠が阻害される可能性があります。
この現象は、温度適応だけでは分布拡大が成功しない理由を説明する新しい概念として注目されています。
種間相互作用への影響
開花時期の変化は、送粉者とのミスマッチを引き起こす可能性があります。Crocus sieberiを用いた研究では、この植物の開花が光周性に強く依存する一方、主要な送粉者であるミツバチの活動開始は温度依存であることが示されています。
気候変動下では温度が先行して上昇するため、光周性に依存する植物と温度依存の送粉者の間で活動時期の同期が崩れる可能性があります。このような表現型の不整合は、個々の種の適応能力だけでなく、生態系全体の機能に影響を及ぼす可能性があります。
まとめ:光周性研究の統合的理解に向けて
野生植物における光周性は、分子レベルの精密な制御機構から、集団の地域適応、さらには生態系の種間相互作用まで、多層的な生物学的意義を持つ現象です。
CO-FTモジュールやフィトクロムを中心とした分子機構は、概日時計と統合されることで、季節という複雑な環境変化を信頼性高く予測する仕組みを実現しています。同一の分子基盤が種や生態的文脈に応じて異なる機能を発揮する柔軟性は、進化の過程で光周性システムが繰り返し改変されてきたことを示しています。
地域適応の観点からは、光周性形質の変異が緯度クラインを形成し、各地域の日長環境への適応を可能にしてきました。しかし、気候変動という急速な環境変化の下では、日長の硬直性が新たな進化的制約となる可能性が浮上しています。
今後の研究では、野生集団における光周性遺伝子の自然変異の包括的な解析、気候変動下での適応的進化の可能性評価、そして生態系レベルでの相互作用ネットワークへの影響予測が重要な課題となるでしょう。光周性という古典的なテーマが、現代の環境問題と交差する地点に、新たな生物学的発見の機会が広がっています。
コメント