脳オルガノイド研究に量子効果の視点が求められる背景
ヒト脳オルガノイドは、iPS細胞や胚性幹細胞から三次元的に分化誘導された「脳様構造体」であり、神経科学・疾患モデル・創薬の文脈で急速に活用が広がっている。近年、この系に「量子生物学」的アプローチを導入しようとする機運が高まっている。その理由は、光合成系やラジカル対スピン化学など他の生体系で量子効果が実証されつつあるなか、神経システムにおける量子現象の役割への関心が増しているからだ。
しかし、単純に「オルガノイドで量子効果を検出する」といっても、何を「量子効果」と定義し、オルガノイドのどの成熟段階でそれが観測可能になるかは、まったく異なる問いである。本記事では、脳オルガノイドの成熟度を多次元的に定義したうえで、量子コヒーレンス・量子もつれ・量子トンネル・量子センシングの各カテゴリとの関係を整理し、現時点での研究ギャップと実行可能な実験設計を示す。
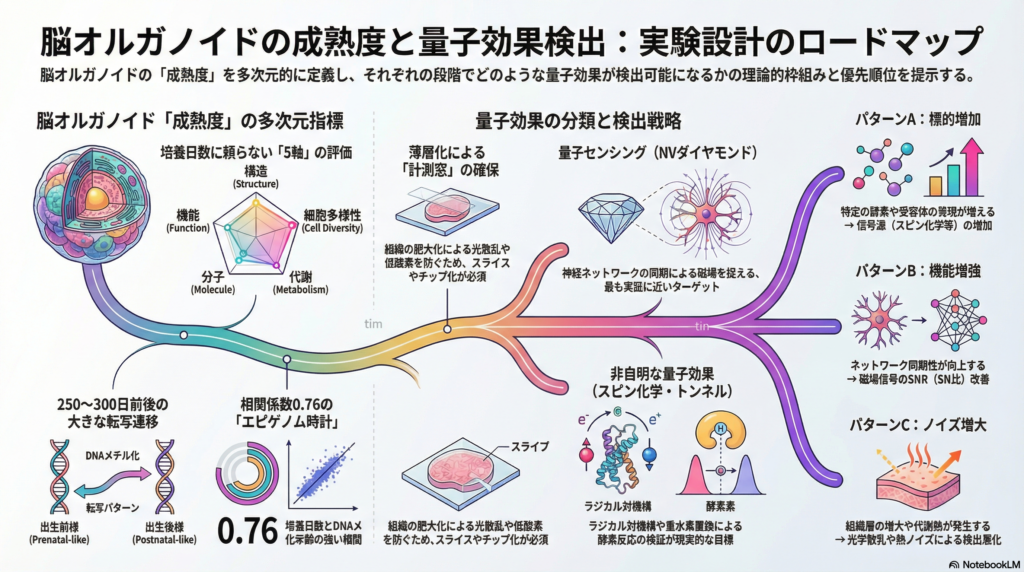
脳オルガノイド「成熟度」の多次元定義
なぜ培養日数だけでは不十分か
多くの研究者が直感的に「培養期間が長いほど成熟している」と考えがちだが、実際には同じ日数でもラインやバッチによって大きなばらつきが生じる。そのため現在の主流は、構造的・細胞多様性・機能・分子・代謝の5軸を組み合わせた多次元評価である。
特に重要な知見として、DNAメチル化年齢と培養日数の間には単調な相関(例として相関係数が0.76程度との報告がある)が見られ、転写署名では培養250〜300日付近に「出生前様から出生後様」への遷移が起こる可能性が示唆されている。この”節目”は、量子計測の時間設計において基準点となりうる。
分子(転写・エピゲノム)指標
シングルセルRNA-seq(scRNA-seq)やスナップショット核RNA-seqを用いると、興奮性・抑制性ニューロン、アストロサイト、オリゴデンドロサイト系列など各細胞種の比率と発達段階マーカーが推定できる。さらに、BrainSpanなど発達トランスクリプトーム参照データベースへのマッピングによって、オルガノイドがどの「胎生週相当」にあるかを推定する枠組みが実装されてきた。
エピゲノム時計(DNAメチル化年齢)は独立した成熟指標として機能する。また、RNA編集モジュールの保存性も脳発達に伴う機能的分子変化を追う指標として活用可能だ。
構造的成熟指標
免疫染色と共焦点顕微鏡による皮質層様マーカーの解析、前後シナプスマーカー、領域同定マーカーを組み合わせることで、組織アーキテクチャの獲得度を定量化できる。超微細構造の評価には電子顕微鏡が用いられ、シナプス後肥厚(PSD)などをナノスケールで確認できる。in vivo移植による成熟促進も有力な手法として知られており、in vitro単独よりも成熟した形態・シナプス・膜特性が形成されうる。
機能(電気生理)指標
多電極アレイ(MEA)を使った長期縦断計測は、自発発火・バースト・ネットワーク同期・局所場電位(LFP)などを追うことができ、成熟に伴う活動複雑化を定量化できる。一部は公開データセットとしても利用可能だ。パッチクランプによる単細胞レベルの膜特性解析(入力抵抗・活動電位閾値・幅・シナプス電流)や、カルシウムイメージングによる広域的な同期パターンの可視化も補完的に用いられる。
代謝・微小環境指標
メタボロミクスや代謝フラックス解析(ATP・乳酸・脂質滴・酸素消費など)は、成熟に伴うエネルギー需要の増大を追うと同時に、組織中心部の壊死・低酸素などの”ノイズ源”を評価する手段でもある。成熟が進むほど酸素拡散の制約が大きくなる点は、後述する量子計測の障害にもなりうる。
量子効果の種類と検出法
量子コヒーレンス:光合成系での実証と神経系への応用の壁
量子コヒーレンスとは、量子状態の位相関係が保持され干渉として観測量に現れる状態を指す。光合成蛋白複合体(FMO複合体など)では、2次元電子分光(2DES)によってフェムト秒〜ピコ秒スケールのコヒーレンス様振動が報告されてきた。ただし、その振動が電子コヒーレンスによるものか振動(vibronic)起源かについては長年の議論があり、「コヒーレンス=機能的量子性」という単純な解釈は修正されてきている。
脳オルガノイドへの適用で最大の壁となるのは、組織の光学散乱・吸収・不均一性である。厚みが増すほどフェムト秒分解の位相情報が失われ、2DES計測が困難になる。現実的な解決策は、オルガノイドを薄層化(スライス・air-liquid interface・on-chip)して光学窓を確保するか、オルガノイド由来の標的分子複合体を単離して計測することになる。
量子もつれとラジカル対スピン化学
生体系における量子もつれは、主にラジカル対機構として議論される。光励起により生成されるスピン相関ラジカル対が外部磁場でスピン進化し、反応収率を変化させるというモデルで、鳥類の磁気受容体として注目されるクリプトクロムがその代表例だ。時間分解EPRや過渡吸収と磁場効果(MFE)計測を組み合わせることで、スピン状態の進行と反応収率変化を定量できる。
理論研究では「温かい系でも数十マイクロ秒以上のスピン量子相関が残り得る」と主張されるが、モデル依存性が高く、ベル不等式違反のような厳密な検証は生体系では困難だ。現時点では「スピン量子ダイナミクスが反応出力を変える」という操作的定義での検出が現実的な目標となる。
量子トンネル:酵素反応の核量子効果
酵素反応における水素(プロトン・ヒドリド)移動では、古典的な障壁透過では説明しきれない反応速度が観測される場合がある。これを量子トンネルとして議論するための主要な実験的証拠は、重水素置換による速度論的同位体効果(KIE)の大きさと温度依存性、および前因子比である。KIEが半古典限界を超える値を示す場合、トンネル寄与が示唆される。
オルガノイドへの適用は、組織全体での直接観測は困難なため、(a)オルガノイドから標的酵素を単離してKIE測定を行う、あるいは(b)同位体置換基質を培地に導入して代謝産物生成速度を高精度定量する、という間接的なアプローチが現実的だ。成熟度の違いが酵素発現・翻訳後修飾・微小環境を変える場合、KIEに差が出る可能性はあるが、現時点での予測は不確実であり、まずは分散推定が必要な段階にある。
量子センシング:生体信号の高感度計測という別カテゴリ
上記3種と性格が異なるが実装上最も重要なのが、量子センサーを使った生体の古典的信号(磁場・温度・常磁性種)の高感度計測だ。これは「非自明な量子効果の存在証明」ではなく、量子力学的な読み出し機構を持つセンサーで生体信号を計測するものである。
NVダイヤモンド(窒素空孔中心)は室温でスピン状態を光学的に読み出せ、磁場・温度・電場に感受性を持つ。単一ニューロンの活動電位(AP)磁場を約10マイクロメートルの近接配置でpT/√Hz級の感度で検出した報告があり、哺乳類組織でも概念実証(PoC)が得られている。原子磁力計(OPM)はfT/√Hz級のノイズフロアを実現し冷却不要のため、将来的なオルガノイドへの近接配置が期待される。また、マイクロ流体チップとNVセンサーを統合する設計が具体化しており、常磁性種検出(T1リラキソメトリ)やNV-NMRがマイクロ流体環境内でデモされている。
成熟度が量子効果検出可能性に影響する3つの経路
現時点では「成熟度指標と量子効果検出感度を同一設計で多時点計測した公開データ」はほぼ存在しない。しかし因果経路は概念的に以下のように整理できる。
パターンA:成熟により標的分子・細胞が出現・増加する。 アストロサイト成熟に伴うNMDA受容体サブユニットの切り替えや特定フラビン系酵素の発現増加が、スピン化学計測の「信号源」を増やす可能性がある。ただし、この因果を実証するには同時計測が必要だ。
パターンB:成熟によりネットワーク活動が増強し、量子センサーで捉えやすくなる。 神経ネットワークの同期性が上がるほど、磁場の有効な双極子モーメントが増し、NV磁力計でのSNRが改善しやすい。この経路は実験的に定義しやすく、最初の実証ターゲットとして最も現実的だ。
パターンC:成熟により光学・熱・磁気ノイズが増え、検出が悪化する。 組織厚の増大、酸素拡散制約による中心壊死、代謝熱・ラジカル生成の増大は、量子センシングにとってノイズ源になりうる。「成熟が進むほど常に有利」とは言えず、薄層化・マイクロ流体統合で計測窓を確保する設計が必須になる。
実験設計:成熟度×量子計測を同時評価するプロトコル
系列A:量子磁力計で成熟依存のネットワーク活動を非接触計測する
最短で実証データを得るための設計として、NVダイヤモンド磁力計とオルガノイドを組み合わせる方法が有望だ。少なくとも4段階の成熟点(例:60日・120日・240日・360日)で評価し、転写署名遷移が示唆される250〜300日付近を高密度に設定する。オルガノイドは薄層化(スライスまたはon-chip)してセンサー近傍(約10マイクロメートル)に配置し、光学散乱と酸素拡散制約を低減する。
主要アウトカムは周波数帯域別の磁場パワー、バースト等の時間ロック平均、マッチドフィルタ後のSNRとする。環境磁場への対策として差動(グラディオメトリ)・磁気シールド・電源ノイズ隔離が必要であり、NV計測に要する高レーザーパワー・マイクロ波が試料に与える熱・光毒性を抑えるためTIR幾何やマイクロ流体設計を採用する。統計的には最初のPoCは各群6〜10サンプルでSNR分布を推定し、次段でパワー再設計する段階的設計が合理的だ。
系列B:スピン化学・トンネル効果を分子現象として検出する
クリプトクロムやフラビン系のラジカル対を対象に、薄層化オルガノイドで磁場掃引(マイクロテスラからミリテスラ)下の過渡吸収・蛍光readoutを取得する設計が現実的だ。NV T1リラキソメトリにより成熟点ごとの常磁性種・ラジカルの時系列マップを取ることで、代謝・酸化還元の成熟依存変化を量子計測の観点で定量できる。酵素トンネルの評価は、成熟点別に標的酵素を精製してKIE測定するか、培地に同位体基質を導入して代謝産物の生成速度を高精度計測する方法が有力だ。
研究ギャップと推奨事項:何から始めるべきか
現状で最大の研究ギャップは、「成熟度指標(独立変数)と量子効果検出感度(従属変数)を同一設計で多時点計測したデータがほぼ存在しない」点に集約される。また、「何をもって量子効果の検出とするか」の操作的定義が研究者間でも揺れており、光合成コヒーレンスの解釈論争がその典型だ。さらに成熟促進(大型化・長期培養)が測定可能性を必ずしも改善せず、むしろ計測窓を悪化させる可能性も念頭に置く必要がある。
推奨される優先順位は以下の通りだ。まず量子センサーで測定する「古典信号」(ネットワーク磁場)を最初の実証ターゲットとし、成熟度×SNRの相関データを作ることが最短経路となる。次に「非自明量子効果」の中では、ラジカル対スピン化学(MFE観測量)と酵素トンネル(KIE)に絞ったPoCを設計する。そして成熟度評価は最初から多次元で取得し、BrainSpan等の参照マッピングを共通スケールとして活用する。最終的には「成熟度データ」「量子計測原データ」「解析コード」を含む公開可能な最小データセット仕様を事前に定めることで、将来のメタ解析への接続性を確保する。
倫理面では、脳オルガノイドの高度化・長期成熟に伴う「意識・苦痛可能性」の問いが継続的な議論対象となっている。現時点で現行培養系が意識・痛覚を持つ生物学的証拠はないとされるが、将来の複雑化に備えた監督体制の整備も並行して進める必要がある。
まとめ:成熟度と量子効果の関係を紐解く二層構造
脳オルガノイド成熟度と量子効果検出可能性の関係は、「量子効果の種類によって構造がまったく異なる」という二層構造で捉えるのが現時点での最も正確な整理だ。
一方では、量子コヒーレンス・もつれ・トンネルといった「非自明な量子効果」をオルガノイドで直接実証した体系的データはまだ乏しく、光学的・実験的課題が大きい。他方では、NVダイヤモンド・原子磁力計・SQUIDなどの量子センサーを使って「古典的生体信号」を高感度に測る統合ロードマップは急速に整備されつつある。
この研究領域で最初に問うべきことは「オルガノイドに量子効果があるか」ではなく、「どの成熟段階で何を、どの観測量として、どれだけの感度で測れるか」という問いの精緻化だ。多次元成熟指標・量子センサー統合・同位体実験の3本柱を組み合わせた縦断実験設計が、今後の研究の基盤となっていくだろう。
コメント